
기억 속의 실험실
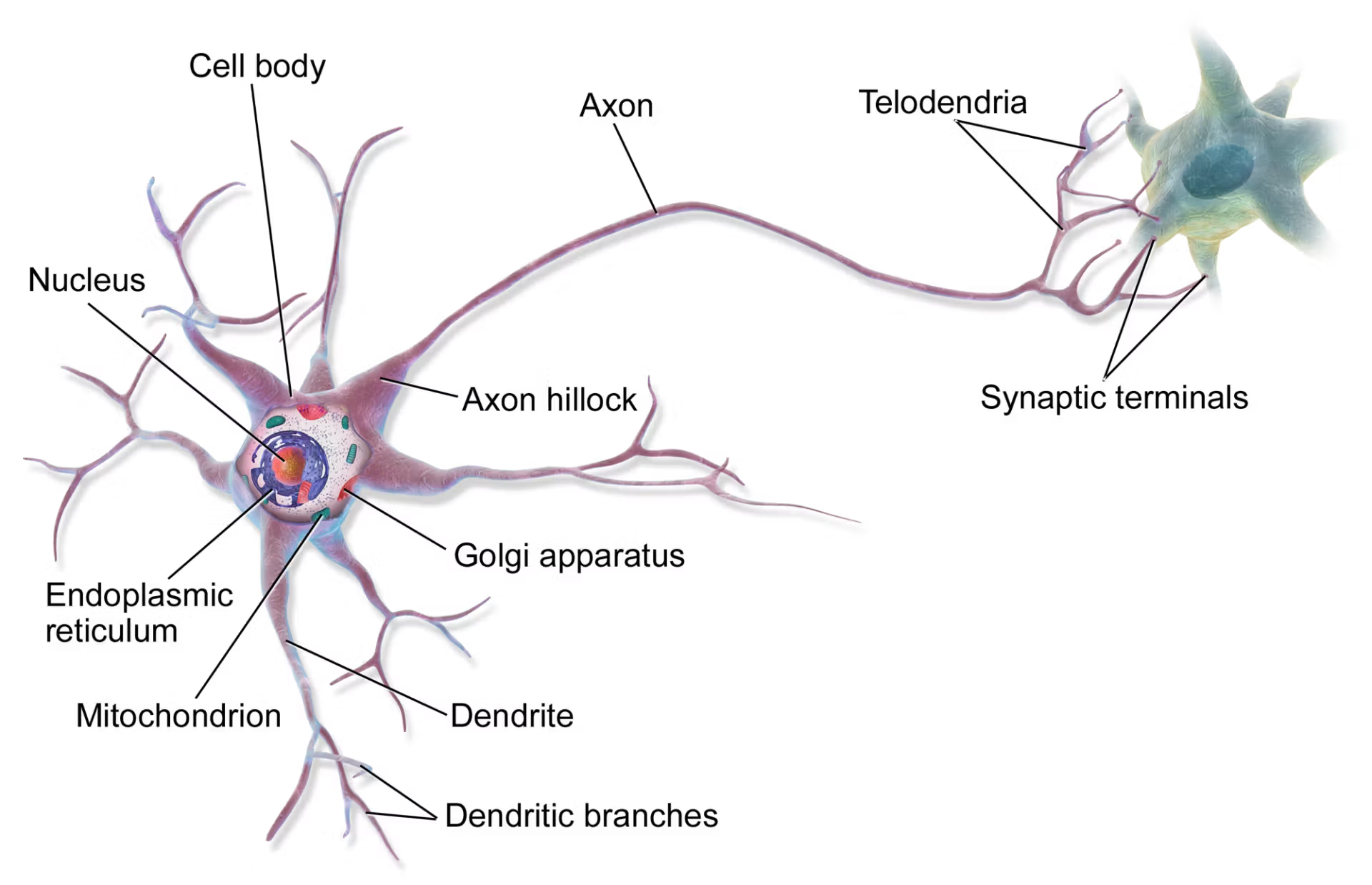
대학원에 들어간 지 얼마 되지 않았을 무렵, 나는 포스텍 생명과학과의 한 작은 실험실에서 리보솜과 처음으로 진지하게 마주쳤다. 내가 맡은 일은 염증 반응과 번역 조절 사이의 관계를 파헤치는 것이었다. 세포가 스트레스를 받으면 어떤 일이 일어나는가. 번역, 즉 mRNA를 단백질로 읽어내는 과정이 멈추면 세포는 무엇을 하는가. 리보솜은 침묵하고, 세포 안에는 번역되지 못한 mRNA들이 뭉쳐 작은 덩어리를 이루기 시작한다. 우리는 그것을 스트레스 과립(stress granule)이라고 불렀다.

나는 그 과립들을 현미경으로 들여다보며 오랜 시간을 보냈다. 형광으로 표지된 단백질들이 세포질 안에서 점처럼 모여드는 모습은 묘하게 아름다웠다. 그 안에 TRAF2가 갇혀 있었다. TRAF2는 TNF-α라는 염증 신호를 세포 핵 안으로 전달하는 중계자인데, 번역이 멈추자 이 중계자도 함께 스트레스 과립 안으로 격리되었다. 번역의 정지가 염증 신호를 차단하는 것이었다. 이 발견은 2005년 논문으로 발표되었다. 그로부터 두 해 뒤, 나는 항염증 지질 매개체인 15d-PGJ2가 번역 개시 인자 eIF4A를 직접 불활성화하여 번역 과정 자체를 틀어막는다는 것을 보여주는 논문을 EMBO Journal에 냈다.
그 시절 나는 번역 조절을 연구하는 사람이 많지 않다는 것을 어렴풋이 느끼고 있었다. 유전체학의 시대가 막 열리고 있었고, 사람들의 시선은 DNA 서열과 유전자 발현량을 측정하는 쪽으로 쏠리기 시작했다. mRNA를 마이크로어레이로 측정하면 유전자 발현을 알 수 있다는 믿음이 퍼져나갔다. 번역을 연구한다는 것은, 조금 낡은 일처럼 보이기 시작했다.
그래서 올해 6월, 카블리 신경과학상 수상자 명단을 보았을 때 나는 묘한 감정을 느꼈다. 기쁨과 안도가 뒤섞인 감정이었다. 크리스틴 홀트, 켈시 마틴, 에린 슈만, 오스왈드 스튜어드. 네 사람이 공동으로 받은 이 상은, 뉴런 안에서 국소적으로 단백질이 합성된다는 사실을 발견하고 증명한 공로에 주어졌다. 번역 조절 연구가, 그것도 가장 정교한 형태의 번역 조절 연구가 과학계 최고 수준의 인정을 받은 것이다. 내가 대학원에서 손으로 만지작거렸던 그 분자들의 이야기가, 결국 이런 결말을 맞이하고 있었다.

그 감정의 뿌리를 따라가다 보면, 단백질 합성이라는 과학의 역사 전체가 펼쳐진다. 이 글은 그 역사에 대한 이야기이자, 뉴런이라는 특별한 세포가 왜 오랜 도그마를 깨뜨리며 우리를 놀라게 했는지에 대한 이야기이다.
리보솜이 태어나던 시절
단백질이 어떻게 만들어지는지를 인류가 이해하기 시작한 것은 1950년대의 일이다. 그 이전까지 단백질 합성은 거의 신비의 영역이었다. 많은 생화학자들은 단백질 합성이 단백질 분해효소의 역반응으로 일어날 것이라고 생각했다. 마치 거꾸로 돌린 분해 기계처럼. 지금 돌이켜보면 황당한 생각이지만, 당시에는 그 대안이 없었다.
알베르 클로드(Albert Claude)는 록펠러 연구소에서 세포를 고속 원심분리기로 갈기갈기 분리하던 중, 미토콘드리아보다 훨씬 작고 RNA가 유달리 풍부한 입자들을 발견했다. 1943년의 일이다. 그는 이 입자들을 마이크로솜(microsomes)이라고 불렀다. 단백질 합성의 실제 무대가 어디인지는 아직 아무도 몰랐지만, 무언가 중요한 것이 그곳에 있다는 예감은 점점 짙어졌다.
폴 자메크닉(Paul Zamecnik)은 1952년에 결정적인 실험을 해냈다. 쥐의 간에서 추출한 세포 내용물을 시험관 안에 넣고, 아미노산을 단백질로 합성하는 데 성공한 것이다. 세포 없이도 단백질 합성이 일어날 수 있다는 것을 보인 이 무세포 합성계(cell-free system)는 이후 수십 년간 분자생물학의 핵심 도구가 되었다. 자메크닉과 마론 호글랜드(Mahlon Hoagland)는 뒤이어 아미노산이 단백질로 합성되기 전에 먼저 ATP와 반응하여 활성화되고, 작은 RNA 분자와 결합한다는 것을 밝혔다. 이것이 훗날 tRNA로 불리게 될 분자의 첫 발견이었다.
1955년, 조지 팔라데(George Palade)는 전자현미경으로 세포질을 들여다보다가 작은 입자들이 세포질 전체에 흩어져 있는 것을 발견했다. 이 입자들, 곧 리보솜(ribosome)이 단백질 합성의 실제 현장임이 나중에 밝혀졌다. 팔라데는 이 공로와 세포 소기관 연구 전반에 걸친 업적으로 1974년 노벨 생리의학상을 받았다. 알베르 클로드와 크리스티앙 드 뒤브(Christian de Duve)와 함께.
그런데 리보솜이 어떻게 작동하는지 이해하려면, 정보를 전달하는 매개체의 존재를 알아야 했다. DNA에서 단백질로 가는 길에는 무언가 중간 전달자가 있어야 했다. 그것이 mRNA라는 생각은 1961년에 구체화되었다. 프랑수아 자코브(François Jacob)와 자크 모노(Jacques Monod)는 유전자 조절을 연구하다가 불안정한 정보 전달자의 존재를 제안했고, 시드니 브레너(Sydney Brenner)와 자코브, 매슈 메설슨(Matthew Meselson)은 같은 해 이 mRNA를 실험적으로 포착해냈다. 세포는 DNA에서 mRNA를 만들고, 리보솜이 mRNA를 읽어 단백질을 합성한다. 분자생물학의 중심 교리(central dogma)가 그 윤곽을 드러낸 것이다.
하지만 유전암호는 아직 해독되지 않았다. DNA나 mRNA의 염기 서열이 어떤 아미노산에 해당하는지 아무도 몰랐다. 이 수수께끼를 풀어낸 것은 마샬 니렌버그(Marshall Nirenberg)와 하인리히 마타이(Heinrich Matthaei)였다. 1961년 5월 27일, 니렌버그의 실험실에서 새벽 늦게 수행된 실험에서 poly-U RNA, 즉 U가 반복된 합성 RNA를 무세포 합성계에 넣자 페닐알라닌만으로 이루어진 짧은 단백질이 만들어졌다. UUU가 페닐알라닌을 지정한다는 것, 유전암호의 첫 글자가 해독된 순간이었다. 이 결과는 같은 해 모스크바에서 열린 국제생화학회에서 발표되었고, 프랜시스 크릭의 주선으로 약 1,000명의 청중 앞에 서게 된 니렌버그는 일약 과학계의 주목을 받았다. 1966년까지 64개의 코돈이 모두 해독되었고, 니렌버그와 고빈드 코라나(H. Gobind Khorana), 로버트 홀리(Robert Holley)는 1968년 노벨 생리의학상을 공동 수상했다.
번역 조절의 세계
단백질이 어떻게 합성되는지를 알게 되자, 다음 질문은 자연스럽게 그 합성이 어떻게 조절되는지로 옮겨갔다. 세포는 어떤 단백질을 얼마나, 언제, 어디서 만들어야 하는지 정밀하게 결정해야 한다. 그 결정의 상당 부분이 번역 단계에서 이루어진다.
진핵세포 mRNA의 5′ 끝에는 특이한 구조가 붙어 있다. m7GpppN이라는 캡(cap) 구조다. 이 캡을 인식하는 단백질 eIF4E는 1978년 나훔 소넨버그(Nahum Sonenberg)와 동료들이 발견했다. eIF4E는 eIF4G, eIF4A와 함께 eIF4F라는 복합체를 이룬다. eIF4A는 ATP를 소비하며 mRNA의 5’UTR에 형성된 이차구조를 풀어, 리보솜이 mRNA 위에 내려앉아 개시 코돈을 향해 이동할 수 있게 한다. 내가 박사과정에서 다루었던 바로 그 eIF4A다. 항염증 물질 15d-PGJ2는 eIF4A의 264번 시스테인에 직접 달라붙어 이 복합체의 조립을 방해한다. eIF4A가 마비되면 리보솜은 mRNA에 올라타지 못하고, 번역은 멈춘다.
번역 조절의 핵심 교차점 중 하나는 mTOR라는 키나아제다. 인슐린, 성장인자, 영양 상태 등 다양한 신호를 통합하여 4E-BP라는 단백질을 인산화하는데, 인산화된 4E-BP는 eIF4E를 붙잡지 못하게 되어 번역이 활성화된다. 반대로 mTOR가 억제되면 4E-BP가 eIF4E를 꽉 붙잡아 번역 개시를 차단한다. mTOR 신호전달계는 암, 노화, 신경 발달 등 거의 모든 세포생물학의 교차로에 있다. 그리고 곧 살펴보겠지만, 시냅스 가소성과 기억의 분자적 기반에도 깊숙이 자리잡고 있다.
1990년대 초반까지 번역 조절 연구는 분자생물학의 한 축이었다. 그런데 1990년대 말부터 유전체 서열 분석과 마이크로어레이 기술이 폭발적으로 발전하면서, 연구의 무게 중심이 전사 조절 쪽으로 크게 기울어지기 시작했다. mRNA의 양을 측정하면 유전자 발현을 알 수 있다는, 편리하지만 근본적으로 불완전한 믿음이 퍼졌다. 번역 조절이나 단백질 분해에 의해 실제 단백질 양이 얼마나 달라지는지는 종종 외면받았다. 초파리 유전학자인 나 역시, 전사 조절 연구가 넘쳐나는 그 시대에 번역 조절을 들여다보는 것이 다소 비주류적인 선택이라고 느꼈다.
물론 이 직관이 완전히 틀린 것은 아니었다. 많은 경우 mRNA 수준은 단백질 양과 상관관계를 갖는다. 그러나 얼마나 강한 상관관계인가에 대해서는 연구자마다 결론이 달랐다. 2011년 슈반호이서(Schwanhäusser) 등은 mRNA가 단백질 양 변이의 약 40%만을 설명한다며 단백질 양은 주로 번역 수준에서 조절된다고 주장했다. 반면 리(Li) 등은 2014년에 측정 오차를 보정하면 mRNA가 최소 56~81%를 설명한다고 반박했다. 어느 쪽이 맞든, 번역의 독립적 기여가 무시할 수 없는 수준임은 분명했다. 2009년 인골리아(Ingolia)가 리보솜 프로파일링(ribosome profiling)이라는 기술을 개발하며 번역이 일어나는 위치와 속도를 전 게놈 수준에서 측정할 수 있게 되었을 때, 번역 연구는 조용히 부흥을 시작했다.
수상돌기에서 폴리리보솜을 보다
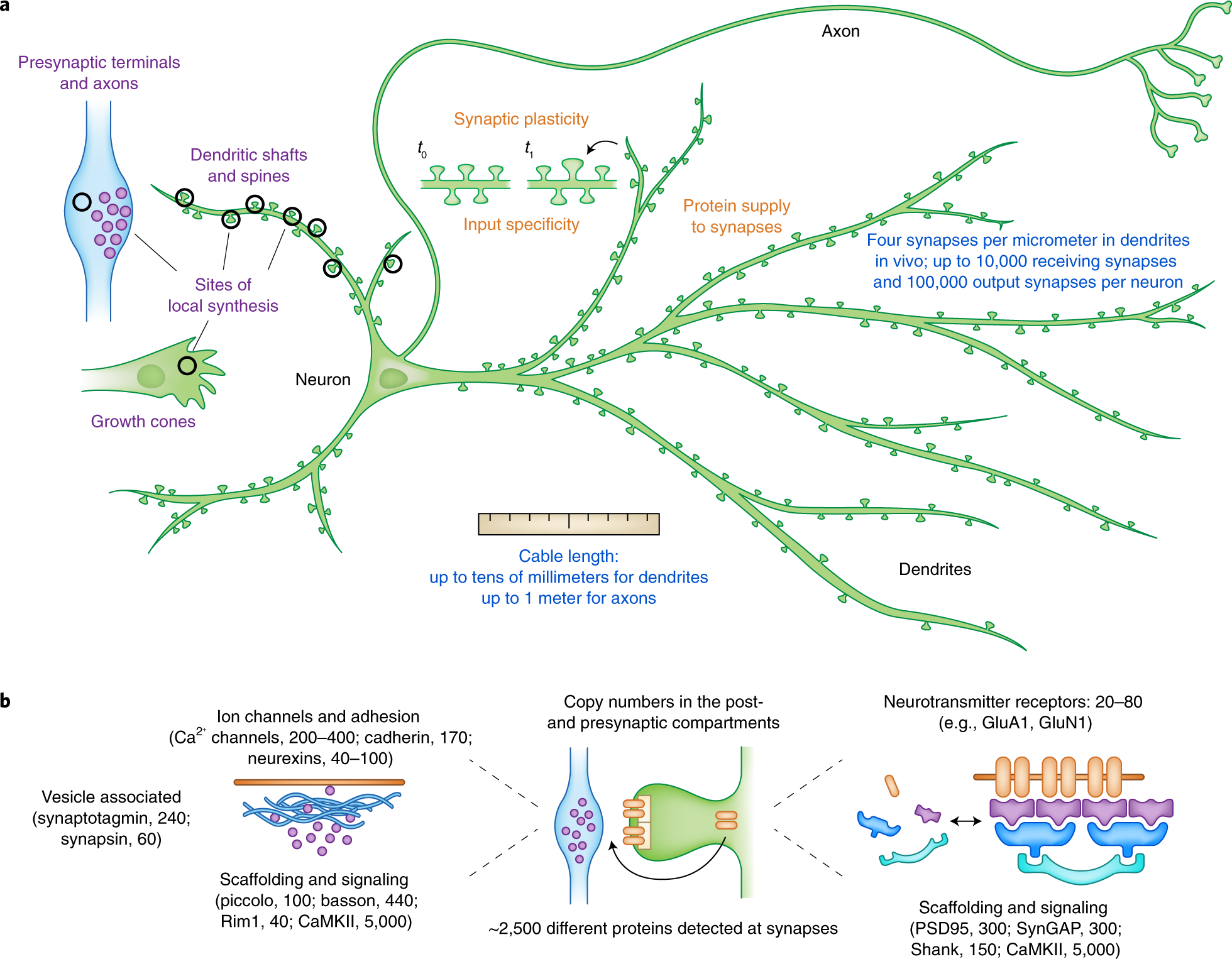
뉴런은 이상한 세포다. 단일 핵을 가지면서도, 수천 개의 시냅스를 형성하는 복잡한 수지상 구조를 뻗어낸다. 인간의 척수 운동 뉴런의 축삭은 1미터에 달하기도 한다. 이 길고 가느다란 돌기들의 끝에서 일어나는 시냅스 전달은, 우리가 기억하고 학습하고 감각하고 운동하는 모든 것의 기초다.
전통적인 세포생물학의 관점에서 단백질 합성은 핵 근처의 세포체에서 일어난다는 것이 정설이었다. 리보솜은 세포체에 있고, 필요한 단백질은 축삭이나 수상돌기로 운반된다는 것. 뇌의 한 뉴런에서 수상돌기 끝까지 단백질을 운반하려면 수 분에서 수 시간이 걸릴 수 있다. 그런데 시냅스 강도의 변화, 즉 학습과 기억은 빠르게 일어난다. 이 시간적 불일치는 오랫동안 크게 문제시되지 않았다. 정설은 강했고, 예외를 상상하는 것은 어려웠다.
오스왈드 스튜어드(Oswald Steward)가 그 정설에 의문을 품기 시작한 것은 1970년대 말이었다. 그는 쥐의 치상회(dentate gyrus) 신경망이 손상된 후 어떻게 회복되는지를 연구하고 있었다. 손상된 부위에 새로운 시냅스가 형성될 때 단백질 합성이 어디서 일어나는지 추적하던 중, 그는 예상치 못한 곳에서 폴리리보솜 신호를 포착했다. 세포체가 아니었다.
1982년, 스튜어드와 윌리엄 레비(William Levy)는 Journal of Neuroscience에 결정적인 논문을 발표했다. 전자현미경으로 치상회 과립세포의 수상돌기 가시(spine)를 조밀하게 절편하여 분석한 결과, 폴리리보솜이 수상돌기 가시의 기저부, 정확히는 가시 목과 수상돌기 줄기가 만나는 지점 바로 아래에 선택적으로 존재한다는 것을 보였다. 단순한 관찰이 아니었다. 정량적 분석에서도 폴리리보솜이 그 특정 위치에 통계적으로 유의미하게 농축되어 있었다. 리보솜이 시냅스 바로 아래에 자리 잡고 있다면, 그것은 거기서 단백질을 합성하기 위해서가 아닌가.
스튜어드는 이후 수년 동안 이 발견의 의미를 추적했다. 그는 재신경지배(reinnervation) 과정에서 이 폴리리보솜의 위치가 변한다는 것을 보여주었다. 시냅스가 형성되는 과정에 국소 번역이 관여한다는 증거였다. 하지만 학계의 반응은 싸늘했다. 단백질 합성은 세포체에서만 일어난다는 통념이 너무 견고했다. 정설에 의문을 제기하는 사람은 항상 힘든 시간을 보낸다.
시냅스 강화와 국소 번역
에린 슈만(Erin Schuman)이 이 문제에 들어온 것은 1990년대 초였다. 슈만은 해마 신경세포에서 장기 강화(LTP, long-term potentiation)를 연구하고 있었다. LTP는 시냅스가 반복적인 자극을 받을 때 강도가 오래 유지되는 현상으로, 기억의 세포생물학적 기반으로 여겨진다.
LTP에는 두 단계가 있다. 초기 단계(E-LTP)는 이미 존재하는 단백질의 변형으로 빠르게 일어나지만, 후기 단계(L-LTP)는 새로운 단백질의 합성을 필요로 한다. 단백질 합성 억제제를 처리하면 초기 LTP는 남지만 후기 LTP가 사라진다는 것이 알려져 있었다. 그렇다면 그 단백질 합성은 어디서 일어나는가.
1996년, 강 혜진(Hyejin Kang)1과 슈만은 Science에 발표한 논문에서 해마 신경섬유망(neuropil) 절편, 즉 세포체를 물리적으로 제거한 후 남은 수상돌기와 축삭 네트워크만 가지고 실험을 수행했다. 세포체가 없으니 번역이 일어날 수 없다는 것이 정설의 예측이었다. 하지만 BDNF와 NT-3 같은 신경영양인자를 가하자, 세포체가 없는 이 절편에서도 즉각적인 단백질 합성과 함께 시냅스 강화가 일어났다. 번역이 세포체가 아닌 수상돌기에서 일어나고 있다는 직접적인 증거였다.

슈만 팀은 여기서 멈추지 않았다. 2001년, 아카루(Aakalu) 등은 CaMKIIα의 조절 서열(5’UTR과 3’UTR)에 불안정한 GFP 리포터를 연결한 구조물을 만들었다. 세포체에서 수상돌기를 분리한 조건에서 BDNF를 처리하자, GFP 형광이 수상돌기의 특정 지점들에서 국소적으로 증가했다. 번역이 일어나고 있었다. 시냅스 근처에 번역이 반복되는 “번역의 핫스팟(translational hot spots)”이 존재한다는 것이 실시간 영상으로 포착된 것이다.

축삭 끝의 자율성 — 크리스틴 홀트의 성장원뿔
수상돌기만이 문제가 아니었다. 개발 중인 신경계에서, 아직 연결을 탐색 중인 어린 신경세포의 축삭 끝에도 같은 질문이 있었다. 성장원뿔(growth cone)은 축삭의 끝에 있는, 주변을 탐지하며 방향을 결정하는 구조다. 화학 유인제가 있으면 그쪽으로 향하고, 반발 신호가 있으면 돌아선다. 이 복잡한 결정이 신경세포 전체의 핵에서 보내는 신호만으로 이루어진다면, 반응은 너무 느릴 것이다.
크리스틴 홀트(Christine Holt)는 1980년대 후반 제노푸스(Xenopus) 개구리 배아에서 이 문제를 직접 검증했다. 그녀는 축삭이 성장 중인 신경세포의 세포체를 잘라내고 축삭 끝의 성장원뿔만 남긴 뒤 관찰했다. 성장원뿔은 정상적으로 발달하고 있었다. 세포체 없이도, 즉 핵에서 오는 신호 없이도 성장원뿔은 스스로 작동할 수 있었다. 필요한 단백질이 세포체에서 만들어져 오는 것이 아니라는 의미였다.
2001년에는 캠벨(Campbell)과 홀트가 네트린-1(netrin-1)에 의한 축삭 유도가 국소 단백질 합성에 의존한다는 것을 보였다. 네트린-1은 성장원뿔이 특정 방향으로 이동하도록 유도하는 화학물질인데, 단백질 합성 억제제를 처리하면 이 유도가 차단되었다. 2006년에는 성장원뿔이 네트린-1 농도 구배를 감지할 때, β-actin mRNA가 성장원뿔의 비대칭적인 위치에서 번역되어 유인성 방향전환을 일으킨다는 것이 밝혀졌다. mRNA가 성장원뿔 안에서 국소화되고, 필요한 순간에 필요한 위치에서 번역된다는 것이었다.
켈시 마틴과 시냅스 특이적 기억
켈시 마틴(Kelsey Martin)이 선택한 모델 생물은 군소(Aplysia californica)였다. 군소는 신경과학자들이 좋아하는 동물이다. 신경세포가 크고 개수가 적어서 개별 시냅스를 추적하기가 용이하다. 에릭 캔델(Eric Kandel)이 군소로 기억의 세포생물학을 연구하여 2000년 노벨상을 받은 것도 이 동물의 장점 덕분이었다.
마틴은 군소 감각 뉴런의 한 가지(branch)만을 선택적으로 자극하는 실험을 설계했다. 하나의 뉴런에는 여러 가지가 있고, 각 가지는 서로 다른 운동 뉴런과 시냅스를 이룬다. 세로토닌을 반복 처리하여 장기 시냅스 촉진(long-term facilitation)을 유도했는데, 그 강화는 자극받은 가지에만, 즉 해당 시냅스에만 선택적으로 일어났다. 단백질 합성 억제제를 처리하면 이 가지 특이적 장기 촉진이 사라졌다. 다른 가지는 변하지 않았다. 시냅스 하나하나가 독립적으로 국소 단백질 합성을 통해 강도를 조절할 수 있다는 것이 직접적으로 증명된 순간이었다.
1997년에 발표된 이 결과는 기억의 시냅스 특이성에 대한 이해를 근본적으로 바꾸었다. 하나의 핵이 어떻게 수천 개의 시냅스를 각각 독립적으로 조절할 수 있는가. 그 답은 국소 번역이었다. 마틴은 이후 CPEB라는 RNA 결합 단백질이 이 과정에서 프리온 유사 성질을 통해 시냅스에서 활성화되어 안정된 기억 흔적을 만드는 데 관여한다는 것도 밝혔다. 번역 조절이 기억의 물질적 기반이라는 개념이 자리를 잡아가고 있었다.
초파리 난자에서 배운 것들
mRNA가 세포 안에서 이동하고 특정 장소에서만 번역된다는 개념은, 사실 신경생물학보다 발생생물학에서 먼저 명확하게 확립되었다. 초파리 유전학자들은 수십 년 전부터 이 메커니즘을 연구해왔다.
초파리 난자형성 과정에서, bicoid라는 mRNA는 미래의 배아에서 머리 쪽이 될 전방(anterior)으로 이동하여 그곳에 농축된다. 알이 수정되면 bicoid가 그 위치에서 번역되어 단백질 농도구배를 만들고, 이 구배가 배아의 앞-뒤 축을 결정한다. oskar mRNA는 반대로 후방으로 이동하여 생식세포질을 형성하고, nanos mRNA는 후방에서 복부 패턴을 결정한다.
이 mRNA들은 어떻게 이동하는가. 플로랑스 베스(Florence Besse)와 안 에프루시(Anne Ephrussi)가 2008년 정리한 리뷰는 그 메커니즘을 명료하게 제시한다. 핵심은 mRNA의 3’UTR에 있는 “zip code”다. 이 서열을 RNA 결합단백질들이 인식하여, mRNA를 세포골격의 운동단백질과 연결한다. kinesin은 미세소관을 따라 mRNA를 후방으로, dynein은 전방으로 운반한다. 이동 중에 mRNA는 번역이 억제된 상태의 RNA-단백질 복합체(RNP) 형태를 유지한다. 제 위치에 도착한 후에야 억제가 풀리고 번역이 시작된다.
oskar의 경우, 도착 전에는 Bruno라는 단백질이 결합하여 번역을 억제하고, 후극에 도착하면 Orb라는 단백질이 poly(A) 꼬리를 늘려 번역을 활성화한다. mRNA는 이렇게 공간적으로, 시간적으로 정밀하게 조절된다. 내가 초파리를 연구하며 익숙하게 접했던 이 체계는, 사실 신경세포의 국소 번역과 깊은 진화적 뿌리를 공유하고 있다. β-actin mRNA의 zip code가 신경세포에서 수상돌기로의 국소화를 지시하는 방식은, bicoid mRNA가 초파리 난자의 전방으로 이동하는 방식과 본질적으로 동일한 원리다. 진화는 같은 해법을 발생과 학습이라는 두 맥락에서 반복해서 사용했다.
국소 번역은 보편적이다 — 2019년의 혁명
스튜어드의 1982년 발견 이후 30여 년이 흐르는 동안, 수상돌기에서의 국소 번역에 대한 증거는 꾸준히 쌓였다. 하지만 시냅스 전 말단, 즉 신호를 방출하는 쪽에서도 국소 번역이 일어나는지는 논란의 여지가 남아 있었다. 시냅스 전 말단은 수상돌기보다 훨씬 작고, 미토콘드리아가 밀집해 있어 리보솜과 구분하기 어렵다는 기술적 문제가 있었다.
2019년, 에린 슈만 팀의 아샤 하프너(Asha Hafner) 등이 Science에 발표한 논문은 이 논쟁을 기술적 혁신으로 해결했다. 이 연구는 단일 기법에 의존하지 않고 여러 접근법을 조합했다. 팽창현미경(expansion microscopy)으로 시료를 물리적으로 약 3.4배 확대하여 공간 분해능을 높였고, FISH로 poly(A) mRNA와 rRNA를 검출했으며, 퓨로마이신 대사 표지로 활발하게 번역이 일어나는 위치를 표시했다. 또한 형광 활성화 시냅토솜 분류(FASS)로 시냅스 전 말단을 분리한 뒤 RNA 서열 분석을 수행했다.
결과는 명확했다. 흥분성과 억제성 시냅스 전 말단의 75% 이상이 리보솜을 함유했고, 80% 이상이 rRNA를 가지고 있었다. 5분간의 대사 표지 실험에서, 흥분성 시냅스 전 말단의 약 37%, 억제성 시냅스 전 말단의 약 44%에서 활발한 번역이 진행 중이었다. 시냅스 후 구획에서는 약 61%에서 번역이 검출되었다. 시냅스 전후 양쪽 모두에서 국소 번역이 보편적으로 일어나고 있었다. 또한 국소 번역이 자극의 종류에 따라 구획 특이적으로 조절된다는 것도 밝혀졌다. mGluR-LTD를 유도하는 자극은 시냅스 후부에서만 번역을 증가시켰고, BDNF는 전후 모두에서, 엔도카나비노이드 수용체를 자극하는 물질은 억제성 시냅스 전 말단에서만 선택적으로 번역을 촉진했다. 각각의 가소성 형태가 제 구획을 갖는다는 것이다.
분리된 시냅토솜에서 검출된 1만 2천여 개의 전사체 중, 시냅스 전 말단에 농축된 것들 중에는 시냅스 소포 방출 기구의 핵심 구성요소들이 포함되어 있었다. 번역 연장 인자 eEF1A와 번역 개시 인자 eIF2도 농축 전사체 중에 있었다. 시냅스에서 번역 기구 자신이 국소적으로 합성된다는 것은, 번역 조절의 자기 증폭적 특성을 시사한다.
왜 이것이 중요한가 — 기억, 질환, 그리고 진화
뉴런이 왜 이렇게 번거로운 방식을 택했는지 생각해보면, 진화적 논리가 보인다. 하나의 핵이 수천 개의 시냅스를 제어하면서, 각 시냅스가 독립적으로 강도를 조절할 수 있으려면, 세포체에서 완성된 단백질을 배달하는 것만으로는 부족하다. 물리적 거리가 너무 멀고, 시간이 너무 걸린다. 필요한 곳에 mRNA를 미리 배치해두고, 자극을 받았을 때 즉시 번역하는 방식이 훨씬 효율적이다.
에드바르드 모저(Edvard Moser) 위원장은 카블리 상 선정 이유를 설명하며 “그들의 연구는 신경과학에서 오랫동안 거의 정설처럼 여겨져 왔던 것을 완전히 뒤집었다”고 말했다. 틀린 말이 아니다. 그런데 이 전복은 순수한 지적 쾌감 이상의 의미를 갖는다.
취약X증후군(Fragile X syndrome)은 FMRP라는 단백질의 기능 상실로 생긴다. FMRP는 수상돌기에 국소화된 mRNA의 번역을 억제하는 RNA 결합단백질이다. FMRP가 없으면 국소 번역이 과잉 활성화되어 시냅스 가소성이 비정상적이 되고, 자폐 스펙트럼 장애와 지적 장애가 나타난다. 하프너 팀이 발견한 시냅스 전 말단의 농축 전사체 중 62개가 FMRP 표적이었다.
알츠하이머 병에서는 eEF1A와 eIF2의 기능이 손상된다. 국소 번역이 제대로 이루어지지 못하면 시냅스 강도를 유지하지 못하고, 기억이 흐려진다. 번역 개시 인자 eIF2의 인산화를 조절하는 통합 스트레스 반응(integrated stress response) 경로를 억제하면 알츠하이머 모델 동물에서 기억 손실이 회복된다는 연구가 있다. 내가 박사과정에서 스트레스 과립과 씨름하며 관심을 가졌던 그 경로가, 지금 신경퇴행성 질환의 치료 표적으로 진지하게 논의되고 있다.
리보솜과 기억, 그 오랜 이야기의 현재
단백질 합성의 역사를 거슬러 올라가면, 클로드의 마이크로솜에서 팔라데의 리보솜으로, 자메크닉과 호글랜드의 무세포 합성계에서 자코브-모노의 mRNA 개념으로, 니렌버그의 유전암호 해독으로 이어지는 한 세기의 발견들이 있다. 그 위에 eIF4F 복합체의 발견과 mTOR 신호전달계의 규명이 쌓였고, 번역 조절이라는 정교한 언어가 만들어졌다.
그리고 그 안에, 스튜어드가 1982년 전자현미경으로 수상돌기 가시 아래에서 본 폴리리보솜이 있다. 모든 것을 바꾼 그 이미지. 그것이 홀트와 마틴과 슈만의 수십 년에 걸친 연구와 합류하여, 뉴런이라는 세포가 얼마나 분산되고 자율적인 방식으로 작동하는지를 드러냈다.
나는 요즘 초파리의 신경 회로를 연구하면서, 가끔씩 내 박사과정 시절의 실험을 떠올린다. 스트레스 과립 안에 갇힌 TRAF2를, eIF4A의 264번 시스테인에 달라붙은 15d-PGJ2를. 그때 나는 번역 조절이 세포가 세상과 상호작용하는 방식의 핵심이라고 믿었지만, 왜 그것이 중요한지를 신경과학의 언어로 설명할 수 없었다. 지금은 조금 더 잘 안다.
기억이라는 것은 시냅스 강도의 지속적인 변화다. 그 변화는 새로운 단백질의 합성을 요구한다. 그 합성은 핵에서 수백 마이크로미터, 때로는 수십 센티미터 떨어진 시냅스 바로 옆에서 일어난다. 세포는 제 경험을 그 자리에서, 즉각적으로, 단백질로 번역한다. 문자 그대로.
카블리 상 수상자들이 보여준 것은 단순히 새로운 세포생물학적 사실이 아니다. 그것은 기억이 어디에 쓰이는가에 대한 대답이다. 기억은 시냅스마다 조각나 있고, 그 각각의 조각은 리보솜이 만든 단백질로 이루어져 있다. 뉴런의 가장 먼 끝에 있는 작은 공장들이, 우리가 배우고 기억하는 모든 것을 만들어낸다.
번역 조절을 연구하던 젊은 대학원생은 결국 초파리를 찾아갔고, 세포가 단백질로 세상을 기억한다는 생각은 그 사이에 과학계 최고의 인정을 받았다. 그것만으로도, 이 긴 이야기를 다시 읽는 데는 충분한 이유가 된다.

참고 문헌
Kim WJ, Back SH, Kim V, Ryu I, Jang SK. (2005). Sequestration of TRAF2 into stress granules interrupts tumor necrosis factor signaling under stress conditions. Mol Cell Biol, 25(6):2450-2462.
Kim WJ, Kim JH, Jang SK. (2007). Anti-inflammatory lipid mediator 15d-PGJ2 inhibits translation through inactivation of eIF4A. EMBO J, 26(24):5020-5032.
Tahmasebi S, Sonenberg N, Hershey JW, Mathews MB. (2019). Protein synthesis and translational control: a historical perspective. Cold Spring Harb Perspect Biol, 11(9):a035584.
Steward O, Schuman EM. (2001). Protein synthesis at synaptic sites on dendrites. Annu Rev Neurosci, 24:299-325.
Hafner AS, Donlin-Asp PG, Leitch B, Herzog E, Schuman EM. (2019). Local protein synthesis is a ubiquitous feature of neuronal pre- and postsynaptic compartments. Science, 364(6441):eaau3644.
Besse F, Ephrussi A. (2008). Translational control of localized mRNAs: restricting protein synthesis in space and time. Nat Rev Mol Cell Biol, 9(12):971-980.
Rheinberger HJ. (2004). A history of protein biosynthesis and ribosome research. Protein Synthesis and Ribosome Structure: Translating the Genome, 1-51.
- 한국인 강혜진 박사에 대해 인터넷을 찾아보았으나 동명이인으로 보이는 연세대학교 생명공학과 강혜진 교수와는 다른 인물로 보이는데, 2002년 논문 이후 행적을 찾을 수 없었다. ↩︎
You must be logged in to post a comment.