초파리 뇌의 성(性)을 둘러싼 한 세기의 유전학

3령 유충의 림프선을 해부하는 일에는 손끝의 조심성이 필요하다. 배쪽 혈관을 따라 늘어선 이 반투명한 조직은 초파리의 조혈 기관, 말하자면 골수에 해당하는 곳이다. 앞엽의 세 구역—전구세포가 머무는 수질구역, 분화한 혈구가 자리 잡는 피질구역, 그리고 줄기세포 니치처럼 행동하는 후방신호중심(PSC)—을 온전히 남긴 채 떼어내려면 유충을 얼음처럼 차가운 완충액에 담그고 핀셋 끝의 압력을 몇 그램 단위로 다스려야 한다. 나는 오랫동안 이 작업을 해오면서, 지극히 당연하다는 이유로 한 번도 묻지 않은 질문이 있었다. 지금 내 핀셋 아래 놓인 이 유충은 수컷인가, 암컷인가.
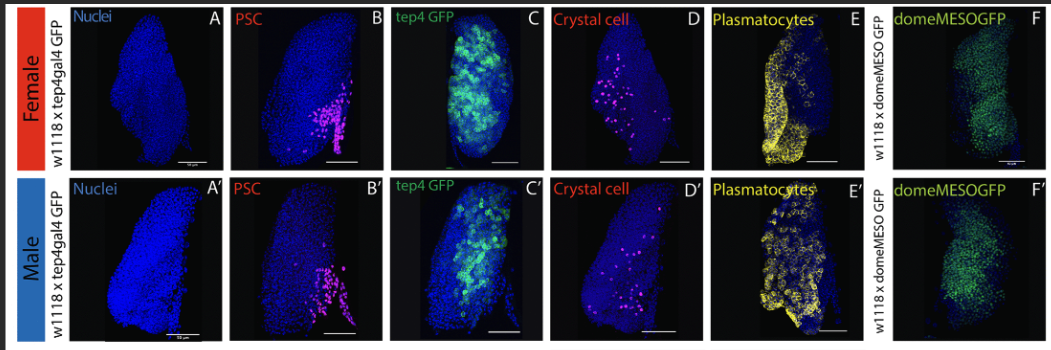
올해 《PLOS Genetics》에 실린 드보스킨과 타넨차프 연구진의 논문은 바로 그 묻지 않았던 질문을 정면으로 파고든다. 브리티시컬럼비아대학의 이 연구는, 몸집이 작은 수컷 유충의 림프선이 암컷의 것보다 작고—수컷은 평균 1,587개, 암컷은 2,183개의 세포로 약 37퍼센트의 차이가 난다—전구세포도 더 적으며 면역세포를 더 적게 만들어낸다는 사실을 보인다. 여기까지라면 그저 몸집 차이의 그림자일 수 있다. 그러나 단일세포 전사체 분석의 결과 앞에서 나는 잠시 손을 멈췄다. 림프선 세포들을 UMAP 위에 펼쳐놓자 지도가 두 개의 반구로 갈라졌고, 그 한쪽을 밝히고 있는 것은 수컷 특이적 긴 비암호화 RNA인 roX1과 roX2였다. 성 결정 유전자 Sex-lethal과 transformer는 암컷 세포에서, msl-2는 수컷 세포에서 켜져 있었다. 혈구가—파리의 백혈구가—제 성별을 기억하고 있었던 것이다. 성이라는 것은 생식소와 생식기에 갇혀 있지 않다. 그것은 피 속으로, 면역 속으로 스며든다. 그리고 피가 그러하다면, 무엇보다 깊은 곳, 뇌 또한 그러할 것이다.
이 지점에서 초파리 행동유전학의 가장 오래되고 가장 근본적인 질문이 되살아난다. 하나의 뇌는 어떻게 수컷이 되는가. 한 마리의 동물은 어떻게 구애하는 존재가 되는가. 이 물음의 계보는 초파리 유전학 자체의 역사만큼이나 길다.
⁂
1915년, 컬럼비아대학 토머스 헌트 모건의 이른바 ‘파리 방’에서, 훗날 최초의 유전자 지도를 그리게 될 앨프리드 스터트반트가 초파리 구애 의식에 관한 최초의 기술을 남긴다. 「초파리에서 성 인식과 성 선택의 문제에 관한 실험」이라는 이 논문에서 그는 수컷의 구애가 일정한 순서를 따르는 정형화된 행동임을 관찰했고, 무엇보다 yellow 돌연변이 수컷이 야생형보다 짝짓기에 덜 성공한다는 사실을 기록했다. 단 하나의 유전자가 행동에 가닿는다는, 그 최초의 암시였다. 흰 눈을 가진 수컷 역시 짝짓기에서 차별받는다는 관찰도 함께였다.
초파리 수컷의 구애는 하나의 고정된 악보처럼 흐른다. 암컷을 향해 몸을 돌리는 지향(orientation), 앞다리로 암컷을 두드리는 태핑, 한쪽 날개를 펼쳐 진동시키며 종 특이적인 ‘사랑 노래’를 연주하는 날개 진동, 암컷의 생식기를 핥는 licking, 그리고 복부를 구부려 시도하는 교미와 실제 교미. 이 일련의 정형행동(fixed action pattern)은 스터트반트에게서 시작해 20세기 중반의 동물행동학자들을 거치며 하나의 완결된 행동목록(ethogram)으로 정리되었다.
그 정리를 완성한 사람이 마거릿 배스톡이다. 옥스퍼드에서 니코 틴베르헌—동물행동학으로 노벨상을 받게 되는—의 지도를 받은 그는 1955년 오브리 매닝과 함께 초파리 구애의 엄밀한 행동목록을 발표했고, 흥분의 수위가 문턱값을 넘을 때마다 다음 행동이 촉발된다는 추동(drive)·문턱 모델을 제시했다. 그리고 이듬해인 1956년, 「행동 양식을 바꾸는 하나의 유전자 돌연변이」라는 제목의 논문에서 그는 다시 yellow로 돌아온다. 배스톡은 yellow 수컷에게서 구애 행동의 요소들이 모두 존재하되 그 활력과 추동이 약해져 있음을 정량적으로 보였다. 단일 유전자 돌연변이가 하나의 행동 양식을 바꿀 수 있음을 처음으로 입증한 이 연구는, 동시에 최초의 본격적인 동물행동학 실험의 하나로 꼽힌다. 틴베르헌에게서 매닝과 배스톡으로 이어지는 이 행동학의 계보는—배스톡은 1959년 매닝과 결혼했다—오랫동안 과학사의 그늘에 묻혀 있다가, 훗날 매슈 콥이 「동물 행동을 바꾼 하나의 유전자 돌연변이: 마거릿 배스톡과 노란 파리」라는 글로 그를 다시 불러냈다.
행동을 향한 유전학의 시선이 특정한 하나의 유전자에 처음으로 이름을 붙인 것은 1963년의 일이다. 쿨비르 길은 수컷 불임 돌연변이를 찾던 중, X선으로 유도한 3번 염색체의 열성 돌연변이 하나를 분리했다. 이 돌연변이의 동형접합 수컷은 교미를 위해 복부를 구부리지 않았고, 다른 수컷을 향해 구애했으며, 수컷들 사이에 구애의 사슬을 촉발했다. 이들은 행동적으로 불임이었다—암컷은 정상이었다. 훗날 이 유전자에는 열매를 맺지 못하는 수컷들의 처지를 따라 fruitless, 곧 ‘열매 없음’이라는 이름이 붙는다. 길이 이를 보고한 두 편의 1963년 기록—《Drosophila Information Service》의 연구 노트와 《American Zoologist》의 초록—이후, 이 돌연변이는 수십 년 동안 계통 보존고 속의 한 기이한 표본으로 남아 있었다.
그것을 하나의 연구 프로그램으로 끌어올린 사람은 브랜다이스대학의 제프리 홀이다. 1970년대와 80년대에 걸쳐 홀과 동료들은 유전적 모자이크—몸의 일부는 수컷, 일부는 암컷인 개체—를 이용해, 구애의 각 단계를 수행하기 위해 뇌의 어느 부위가 수컷이어야 하는지를 지도로 그려냈다. 홀은 또한 후배 학자 참발로스 키리아쿠와 함께 구애 노래를 파고들었다. 1980년 이들이 발표한 결과에 따르면, 수컷의 사랑 노래에서 음 사이 간격(interpulse interval)은 약 1분 주기로 느리게 오르내리는 리듬을 갖는데, 이 리듬이 period 돌연변이에 의해 교란되었다. period는 다름 아니라 홀 자신이 훗날 마이클 로스배시, 마이클 영과 함께 클로닝하여 2017년 노벨 생리의학상을 받게 되는 그 생체시계 유전자다. 구애와 시계는, 이야기의 첫머리에서부터 이미 한 몸으로 얽혀 있었다.

⁂
그리고 1996년이 온다. 이 이야기의 무게중심이자 전환점. 서로 다른 두 연구실이, 독립적으로, 거의 동시에 fruitless를 클로닝했다.
먼저 그해 9월 3일, 야마모토 다이스케 연구진의 이토 히로키 등이 《PNAS》에 논문을 실었다. 이들은 satori—선(禪)의 ‘깨달음’에서 딴 이름으로, 암컷에게 무심한 수컷들의 처지를 빗댄 작명이었다—라 명명한 돌연변이에서 출발했다. 이 논문은 satori가 fruitless와 같은 자리(세포유전학적 위치 91B)의 대립유전자임을 밝히고, 이 유전자가 BTB 도메인과 아연 손가락(zinc finger)을 지닌 전사인자를 암호화함을 보였으며, 돌연변이 수컷에게서 수컷 특이적 근육인 ‘로런스 근육’이 사라진다는 사실을 함께 보고했다. 야마모토는 벤저의 행동 돌연변이 연구에 자극받아 1988년부터 이 satori 스크리닝을 시작했고, 그의 문하에서 이토 히로키, 기무라 겐이치, 고가네자와 마사유키 같은 이름들이 자라났다.
두 달여 뒤인 그해 11월, 브루스 베이커 연구실의 리사 라이너 등이 《Cell》에 논문을 발표했다. 홀, 바버라 테일러, 스티븐 워서먼이 저자로 함께한 이 연구는 워서먼 연구실의 P인자 삽입 대립유전자들에 기대어 fruitless를 클로닝하고, 이 유전자의 성 특이적 스플라이싱이 transformer와 transformer-2에 의해 조절됨을 밝히며, 수컷 특이적 동형단백질 FruM을 규정했다. 인쇄 시점만 놓고 보면 이토 등이 두어 달 앞서지만, 학계는 서로 다른 대립유전자와 서로 다른 전략으로 도달한 이 두 성과를 관행적으로 나란히—’이토 등 1996; 라이너 등 1996’—공동 발견으로 인용한다. 그리고 그 앞에는, 잊지 말아야 할, 1963년의 길이 있다. fruitless를 흔히 한 연구실이 ‘발견했다’고 말하곤 하지만, 정확히는 길이 돌연변이를 발견했고(1963), 두 연구진이 유전자를 독립적으로 클로닝하고 규명했다(1996).
이 유전자의 분자적 논리는 우아하다. fru는 3번 염색체 91B에 놓여 있으며, 네 개의 프로모터(P1–P4)에서 전사되어 5′과 3′ 양쪽 끝에서 광범위한 선택적 스플라이싱을 겪는다. 성 특이적 전사체는 가장 먼 P1 프로모터에서 나온다. 암컷에서는 Tra 단백질과 Tra-2가 이 전사체를 조기 종결코돈이 생기도록 스플라이싱하여, 기능하는 단백질이 만들어지지 않는다. 수컷에서는 기본 스플라이싱이 일어나 FruM 단백질이 만들어진다. FruM은 N말단의 BTB(단백질 상호작용) 도메인을 공유하고, 여러 종류의 아연 손가락 DNA 결합 도메인 가운데 하나를 C말단에 지닌다. 이 단백질은 중추신경계 뉴런의 약 2퍼센트—대략 1,200에서 2,000개 사이로 추정되는—에서 발현된다. 요컨대 fru는 양성 모두에서 전사되지만, 기능하는 Fru 단백질은 오직 수컷에게만 존재하며, 그 수컷 특이적 전사인자가 뇌를 구애에 맞게 배선한다.
이 유전자가 어디에 놓여 있는지를 이해하려면 초파리 성 결정의 위계 전체를 보아야 한다. X염색체와 상염색체의 비율이 암컷에서 Sex-lethal을 켜고, Sxl은 transformer의 스플라이싱을 조절하며, 암컷 특이적 Tra 단백질과 Tra-2가 하위의 두 갈래—doublesex와 fruitless—를 여성형으로 스플라이싱한다. 여기서 개념적으로 결정적인 분업이 성립한다. doublesex는 몸의 성적 분화, 곧 형태와 색소와 생식기와 대부분의 비신경 이형성을 관장하고, fruitless는 성적 행동의 신경적 기질, 곧 수컷의 구애를 관장한다. fru는 구애 행동을 조절하기 위해 중추신경계에서 특이적으로 기능하는 것으로 밝혀진 최초의 유전자다. 그리고—여기서 이야기의 원이 닫히기 시작하는데—Sxl은 동시에 msl-2의 번역을 억제함으로써 유전량 보상(dosage compensation)을 통제한다. 드보스킨의 림프선 데이터에서 켜져 있던 바로 그 roX–msl 축이다. 뇌의 성을 결정하는 위계의 가장 윗부분이, 파리의 피에도 똑같이 새겨져 있었던 것이다.
doublesex를 분자적으로 규명한 것 역시 베이커 연구실이었다. 1989년 커티스 버티스와 브루스 베이커는 dsx가 tra/tra-2의 통제 아래 성 특이적으로 스플라이싱되어 수컷형 DsxM과 암컷형 DsxF를 만들며, 이 둘이 공통의 DNA 결합 영역을 공유하되 C말단에서 갈라진다는 것을 보였다. 이 DNA 결합 영역, 곧 DM 도메인은 예쁜꼬마선충의 mab-3와 공유되며, 포유류에까지 보존된 DMRT 유전자 집안의 시조가 되었다. 생쥐의 Dmrt1은 출생 후 정소 분화에 필수적이고, 조류에서는 DMRT1의 유전량이 성을 결정한다. 성 결정의 메커니즘이 종마다 어지럽게 갈라지는 진화의 풍경 속에서, DM 도메인은 드물게도 깊이 보존된 한 가닥의 실이다.
이 모든 것의 중심에 브루스 베이커(1945–2018)가 있다. 리드 칼리지를 졸업하고 워싱턴대학에서 래리 샌들러의 지도로 박사학위를 받은 뒤 위스콘신에서 제임스 크로의 문하에서 박사후연구를 거친 그는, UCSD와 스탠퍼드를 거쳐 말년에는 하워드휴스의학연구소 재닐리아로 옮겼다. 1980년 베이커와 리지가 발표한 「성과 단일 세포」는 성이 세포 자율적으로 결정된다는 것을 보인 기념비적 연구였다. 흥미롭게도 홀 역시 샌들러의 제자였으니, fruitless를 클로닝한 두 축—베이커와 홀—은 같은 스승에게서 뻗어 나온 두 가지였던 셈이다. 베이커는 2018년 7월 1일, 일흔둘의 나이에 갑작스레 세상을 떠났다.

⁂
2005년은 fruitless의 기적의 해라 불릴 만하다. 취리히에서 에른스트 하펜의 지도로 박사학위를 받고 버클리에서 코리 굿맨의 문하를 거친 오스트레일리아의 신경생물학자 배리 딕슨은, 빈의 연구소에서 fru를 수컷형 혹은 암컷형으로 항상 스플라이싱되도록 조작한 파리를 만들었다. 에브루 데미르와 딕슨의 이 실험 결과는 충격적이었다. 유전적으로 암컷인 개체에게 FruM을 강제로 발현시키자, 그 암컷이 다른 암컷을 향해 구애했다—날개 노래를 포함한 수컷 구애의 전 과정을 펼치면서. 수컷형 fru 스플라이싱은 수컷 구애에 필요할 뿐 아니라, 대체로 충분했다. 같은 해 베이커 연구실의 마놀리 등도 나란히 이 사실에 도달했다. 키리아쿠는 《Nature》의 논평에서 이를 두고 “초파리의 성은 fruitless하다”고 썼다.
같은 해, 야마모토 연구실의 기무라 등은 뇌 속의 성적 이형 뉴런에 이 유전자를 직접 대응시켰다. 이들이 밝힌 mAL 뉴런 무리는 암컷에서 약 5개, 수컷에서 약 30개다. 이 차이를 만드는 것은 성장이 아니라 죽음이었다. 수컷에서는 FruM이 프로그램된 세포사를 억제하여 이 뉴런들을 살려두지만, FruM이 없는 암컷에서는 그 뉴런들이 죽는다. 성적 이형성이 사형집행인의 손끝에서 조각되고 있었던 것이다. 2008년 기무라 등은 여기서 한 걸음 더 나아가 P1 뉴런 무리를 규명했다. fru와 dsx를 함께 발현하는 약 20개의 수컷 특이적 사이뉴런으로, 암컷에서는 여성형 단백질 DsxF의 작용으로 죽어 사라진다. 그런데 이 뉴런들을 인위적으로 활성화하면, 수컷은 구애 의식의 전 레퍼토리를 남김없이 펼친다. 구애의 지휘 본부인 셈이다. fru가 수컷 구애의 ‘잠재력’—신경적 기질—을 세우고, dsx가 그것을 조율하며 함께 필요해지는 이 이중의 논리는, 이후 페로몬 처리 회로(데미르와 딕슨이 리처드 액설과 함께 밝힌, 수컷 페로몬 cVA가 암수의 뇌에서 다르게 처리되는 성적 이형 회로)와 커넥톰 규모의 회로 지도로 확장되어 갔다.
⁂
바로 이 지점에서 이야기는 실험대를 떠나 문화 속으로 들어선다. 수컷 파리가 수컷을 향해 구애하는 순간, 인간의 상상력은 하나의 단어—동성애—와 하나의 유전자를 향해 손을 뻗기 때문이다.
fruitless가 클로닝되기 한 해 전인 1995년, 미국 국립보건원의 장위칭과 워드 오든발트는 성과는 무관한 표지 유전자 mini-white를 과발현시키는 실험을 하던 중, 수컷들이 서로 구애하며 사슬을 이루는 현상과 우연히 마주쳤다. 그해 6월 《Time》은 이를 “동성애 유전자를 찾아서“라는 제목으로 대서특필했다. 경악한 오든발트는 《New York Review of Books》에 반박문을 실어, 자신들은 결코 인간과의 그런 유비를 끌어낸 적이 없으며 실험 결과를 동성애에 관한 선입견에 끼워 맞춘 적도 없다고 못박았다. 훗날 2008년의 genderblind 연구는 신경교세포의 글루탐산 수송체를 다루는데, 이 유전자를 잃은 수컷은 다른 수컷을 향해 구애하지만—그 행동은 성체에서 몇 시간 안에 약물로 껐다 켤 수 있었다. 도파민을 높여도(2008년 류 등), 강한 빛을 쬐어도 야생형 수컷에게서 같은 행동이 나타난다.
여기에 이 이야기의 핵심이 있다. 파리에서 ‘성적 지향’이란 표적 인식과 감각 관문의 문제, 곧 하드와이어된 운동 프로그램이 어느 방향을 향하는가의 문제다. 그것은 정체성도, 욕망도, 인간의 그것도 아니다. fruitless를 ‘동성애 유전자’라 부르는 것은 단지 틀린 것이 아니라 범주 오류이며, 그 오류는 과학에도, 그 은유가 투사된 사람들에게도 실질적인 해를 끼쳤다. 파리는 하나의 신경계가 어떻게 구애 행동의 잠재력을 짓는지를 우리에게 말해줄 수 있다. 그러나 파리는, 한 남자가 다른 남자를 사랑한다는 것이 무엇인지에 대해서는 아무것도 말해줄 수 없다.

⁂
다시 림프선으로 돌아온다. 드보스킨의 연구는, 이 한 세기의 역사가 벼려낸 바로 그 도구—transformer를 이용한 여성화—로 조직 특이적으로 성적 정체성을 바꾸어가며, 성차가 피와 면역에까지 가닿을 뿐 아니라 결코 하나의 얼굴이 아님을 보여준다. 어떤 형질은 몸집을 따라가고, 어떤 형질은—결정세포(crystal cell)처럼—몸집을 넘어선다. 어떤 차이는 니치가, 어떤 차이는 인슐린 신호가 매개한다. 어떤 차이는 크기로 보정하면 사라지고, 어떤 차이는 끝내 남는다. 성에는 단일한 주스위치가 없다. 사랑에 단일한 유전자가 없듯이. fru는 스위치가 아니라 잠재력의 건축가이며, 그 잠재력은 오직 맥락의 바다—dsx, 페로몬, 경험, 동물 전체—속에서만 실현된다.
스터트반트가 노란 수컷 한 마리가 짝을 얻지 못하는 모습을 지켜보던 1915년에 시작되어, 하나의 스플라이싱이 뇌가 구애할지를 결정한다는 발견으로 1990년대에 그 깊이에 이른 이 한 세기는, 단일 유전자의 승리가 아니라 그 겸허함으로 끝난다. 우리가 배운 것은 성이 하나의 유전자에 쓰여 있다는 것이 아니라, 성이 도처에 쓰여 있다는 것이다—뇌에도 피에도, 살아남는 뉴런에도 죽도록 만들어진 뉴런에도. 그리고 그것을 읽어내는 일은, 우리 과학의 가장 오래된 유혹을 견디는 데서 시작된다. 이름을 설명으로, 유전자를 운명으로 착각하려는 그 유혹을.

참고문헌
초기 역사와 동물행동학
- Sturtevant, A. H. (1915). Experiments on sex recognition and the problem of sexual selection in Drosophila. Journal of Animal Behavior, 5, 351–366.
- Bastock, M., & Manning, A. (1955). The courtship of Drosophila melanogaster. Behaviour, 8, 85–111.
- Bastock, M. (1956). A gene mutation which changes a behaviour pattern. Evolution, 10, 421–439.
- Spieth, H. T. (1974). Courtship behavior in Drosophila. Annual Review of Entomology, 19, 385–405.
- Cobb, M. (2007). A gene mutation which changed animal behaviour: Margaret Bastock and the yellow fly. Animal Behaviour, 74, 163–169.
fruitless 돌연변이와 클로닝
- Gill, K. S. (1963). A mutation causing abnormal courtship and mating behavior in males of Drosophila melanogaster. American Zoologist, 3, 507 (초록); Drosophila Information Service (연구 노트).
- Hall, J. C. (1977). Portions of the central nervous system controlling reproductive behavior in Drosophila melanogaster. Behavior Genetics, 7, 291–312.
- Kyriacou, C. P., & Hall, J. C. (1980). Circadian rhythm mutations in Drosophila melanogaster affect short-term fluctuations in the male’s courtship song. PNAS, 77(11), 6729–6733.
- Ito, H., Fujitani, K., Usui, K., Shimizu-Nishikawa, K., Tanaka, S., & Yamamoto, D. (1996). Sexual orientation in Drosophila is altered by the satori mutation in the sex-determination gene fruitless that encodes a zinc finger protein with a BTB domain. PNAS, 93(18), 9687–9692.
- Ryner, L. C., Goodwin, S. F., Castrillon, D. H., Anand, A., Villella, A., Baker, B. S., Hall, J. C., Taylor, B. J., & Wasserman, S. A. (1996). Control of male sexual behavior and sexual orientation in Drosophila by the fruitless gene. Cell, 87(7), 1079–1089.
성 결정 위계와 doublesex / DMRT
- Baker, B. S., & Ridge, K. A. (1980). Sex and the single cell. I. On the action of major loci affecting sex determination in Drosophila melanogaster. Genetics, 94(2), 383–423.
- Burtis, K. C., & Baker, B. S. (1989). Drosophila doublesex gene controls somatic sexual differentiation by producing alternatively spliced mRNAs encoding related sex-specific polypeptides. Cell, 56(6), 997–1010.
- Raymond, C. S., Murphy, M. W., O’Sullivan, M. G., Bardwell, V. J., & Zarkower, D. (2000). Dmrt1, a gene related to worm and fly sexual regulators, is required for mammalian testis differentiation. Genes & Development, 14(20), 2587–2595.
신경회로와 성적 이형성 (2005–현재)
- Demir, E., & Dickson, B. J. (2005). fruitless splicing specifies male courtship behavior in Drosophila. Cell, 121(5), 785–794.
- Stockinger, P., Kvitsiani, D., Rotkopf, S., Tirián, L., & Dickson, B. J. (2005). Neural circuitry that governs Drosophila male courtship behavior. Cell, 121(5), 795–807.
- Manoli, D. S., Foss, M., Villella, A., Taylor, B. J., Hall, J. C., & Baker, B. S. (2005). Male-specific fruitless specifies the neural substrates of Drosophila courtship behaviour. Nature, 436(7049), 395–400.
- Kimura, K., Ote, M., Tazawa, T., & Yamamoto, D. (2005). Fruitless specifies sexually dimorphic neural circuitry in the Drosophila brain. Nature, 438(7065), 229–233.
- Kimura, K., Hachiya, T., Koganezawa, M., Tazawa, T., & Yamamoto, D. (2008). Fruitless and Doublesex coordinate to generate male-specific neurons that can initiate courtship. Neuron, 59(5), 759–769.
- Datta, S. R., Vasconcelos, M. L., Ruta, V., et al. (2008). The Drosophila pheromone cVA activates a sexually dimorphic neural circuit. Nature, 452(7186), 473–477.
- Rideout, E. J., Dornan, A. J., Neville, M. C., Eadie, S., & Goodwin, S. F. (2010). Control of sexual differentiation and behavior by the doublesex gene in Drosophila melanogaster. Nature Neuroscience, 13(4), 458–466.
‘동성애 유전자’ 논쟁
- Zhang, S.-D., & Odenwald, W. F. (1995). Misexpression of the white (w) gene triggers male-male courtship in Drosophila. PNAS, 92(12), 5525–5529.
- Odenwald, W. F. 외 (1995년 11월 2일). Genes and Sexuality: An Exchange. The New York Review of Books.
- Grosjean, Y., Grillet, M., Augustin, H., Ferveur, J.-F., & Featherstone, D. E. (2008). A glial amino-acid transporter controls synapse strength and courtship in Drosophila. Nature Neuroscience, 11(1), 54–61.
리뷰 및 관련 문헌
- Yamamoto, D., & Koganezawa, M. (2013). Genes and circuits of courtship behaviour in Drosophila males. Nature Reviews Neuroscience, 14(10), 681–692.
- Millington, J. W., & Rideout, E. J. (2018). Sex differences in Drosophila development and physiology. Current Opinion in Physiology, 6, 46–56.
- Rideout, E. J., Narsaiya, M. S., & Grewal, S. S. (2015). The sex determination gene transformer regulates male-female differences in Drosophila body size. PLoS Genetics, 11(12), e1005683.
도입부 논문
- Dvoskin, A., Ho, K. Y. L., Allara, M., Janz, N., Rideout, E., Girard, J. R., & Tanentzapf, G. (2026). Sex differences in the regulation and function of cellular immunity in Drosophila. PLOS Genetics, 22(7), e1012151.